骨髄由来細胞のNOX1/NADPHオキシダーゼが腸管バリア機能を調節する
LPSにより回腸でNOX1由来活性酸素種の産生が亢進
クローン病や潰瘍性大腸炎などの炎症性腸疾患(IBD)の発症や進展には、活性酸素種(ROS)や一酸化窒素(NO)による腸管バリア機能障害が関与することが知られています。
しかし、腸管バリア機能障害に関与するROSの産生源や作用機構については明らかではありません。
我々はNOX1欠損マウスを用い腸管バリア機能障害におけるNOX1の役割を検討しました。
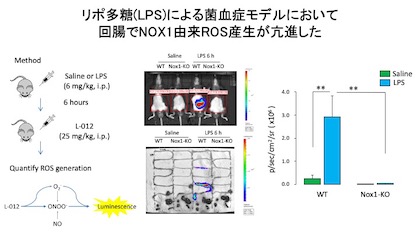
リポ多糖(LPS)によるマウス菌血症モデルにおいて,ROS産生を in vivo imaging system (IVIS)を用い非侵襲的に測定したところ,
野生型マウス(WT)の腹部において著明な発光シグナル(ROS産生)の増加が認められましたが,NOX1欠損マウス(NOX1-KO)ではほとんど発光が認められませんでした.
また,摘出臓器でROS産生を測定したところ、LPSにより回腸で特に強いNOX1由来ROS産生を認めました.
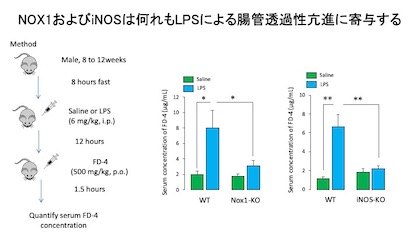
続いてLPSによる腸管バリア機能障害における,NOX1由来活性酸素種(ROS)および一酸化窒素(NO)の関与を検討しました。
腸管バリア機能障害は、経口投与した蛍光標識デキストランFD-4の血中への移行量を指標として検討しました。
LPSを投与した野生型マウス(WT)では血清FD-4濃度が有意に増加しましたが、NOX1欠損マウス(NOX1-KO)およびiNOS欠損マウス(iNOS-KO)では有意に抑制されました.
このことからNOX1およびiNOSは何れもLPSによる腸管透過性亢進(腸管粘膜バリア機能障害)に寄与すると考えられます。
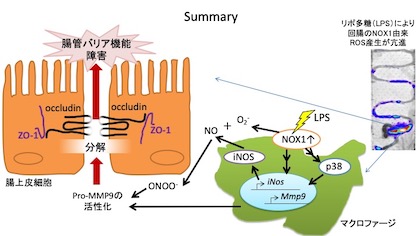
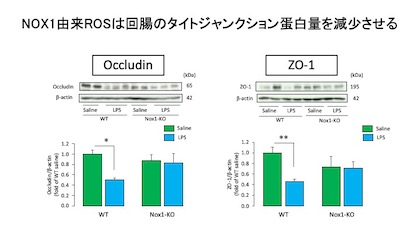
腸管バリア機能の維持にはOccludinやZonula occludens protein 1(ZO-1)などのタイトジャンクション蛋白が重要な役割を果たしています。
LPSを投与するとWTの回腸ではOccludinおよびZO-1タンパクが有意に減少しましたが、NOX1-KOでは変化は認められませんでした。
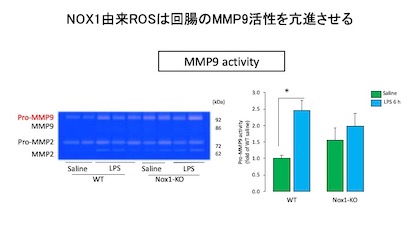
Occludinはマトリックスメタロプロテアーゼ(MMPs)により分解されることが知られています。
そこでマウス回腸組織のMMPs活性をゼラチンザイモグラフィーで測定しました。
LPSを投与したWTではpro-MMP9活性の上昇が認められましたが、NOX1-KOでは変化が認められませんでした。
骨髄由来細胞のNOX1がLPSによる腸管透過性亢進に寄与
我々の研究室ではLPSにより腸上皮でNOX1蛋白の発現が上昇することを見出しています。
一方,MMPsは主にマクロファージや好中球等の骨髄由来細胞から分泌されることが知られています。
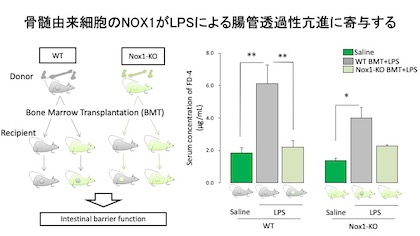
腸管バリア機能の障害に関与するNOX1由来ROSの産生源を明らかにするために,骨髄移植実験を行いました。
WTにNOX1-KOの骨髄細胞を移植したマウスではLPSによる腸管透過性の亢進が有意に抑制されました。
対して,NOX1-KOにWT由来骨髄細胞を移植したマウスでは、LPSによる腸管透過性の亢進が認められました。
以上の結果から骨髄由来細胞のNOX1がLPSによる腸管透過性亢進に寄与することが明らかになりました。
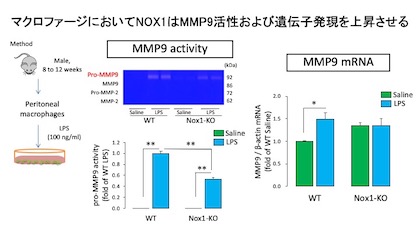
続いてマウス腹腔マクロファージを単離し,LPSを処置したところWTではpro-MMP9の活性が著明に上昇しましたが、NOX1-KO由来のマクロファージでは有意に抑制されました。
またLPSを処置した野生型マクロファージでMMP9のmRNAが有意に増加しましたが,NOX1-KOでは差異が認められませんでした。
以上の結果から、菌血症において骨髄由来細胞のNOX1がMMP9の発現上昇および活性化により,腸上皮細胞のタイトジャンクション蛋白を分解し、腸管バリア機能障害を引き起こすことが明らかとなりました。